Зябровий тип дихання характерний для. Філогенез органів дихання загальна характеристика типів дихання. Клас Земноводні = Амфібії
Зябровий апарат у хордових еволюціонував у напрямі утворення зябрових пелюсток. Зокрема, у риб розвинулося 4-7 зябрових мішків, які є щілинами між зябровими дужками і містять велика кількістьпелюсток, які пронизані капілярами (рис. 190). У риб у диханні бере участь також повітряний міхур.
Зяброве дихання є типовим водним диханням. Фізіологічне призначення зябер - постачання організму киснем. Вони передають кисень із зовнішнього середовища в кров.
Шкірне дихання як найпервинніше у філогенезі і онтогенезі потім змінюється спеціальним, зябровим диханням, але все ж таки відому роль продовжує грати до кінця життя риби.
Органи дихання. Зябра суть органи дихання. Вони лежать по обидва боки голови. Основою їх служать зяброві дуги. У більшості випадків у наших прісноводних риб, за винятком тільки мініг, зябра покриті зовні кришками, і порожнина їх повідомляється з порожниною рота. На зябрових дугах розташовані дворядні зяброві пластинки. Кожна зяброва платівка довгастої, загостреної, мовоподібної форми, має в основі своєму хрящувату тичинку, укладену в кістяний чохлик і досягає до її вільного кінця. Уздовж внутрішнього краю зябрової пластинки проходить гілочка зябрової артерії, що приносить венозну кров, а вздовж зовнішнього краю - гілочка зябрової вени, що відводить артеріальну кров. Від них відходять волосяні судини. По обидва плоскі сторони зябрової пластинки знаходяться листоподібні пластинки, які і служать власне для дихання або обміну газів. Якщо на зябрової дузі знаходиться лише один ряд пластинок, то така називається напівжаброю.
У бичків дихання у вологому повітрі забезпечується шкірою голови, ротовою та зябрової порожниною. Слизова оболонка цих порожнин добре постачається кровоносними судинами. Повітря забирається ротом, кисень поглинається в ротовій або зябрової порожнини, а газ, що залишився, викидається назад через рот. Цікаво, що багато бичків немає плавального міхура, й у повітряного дихання пристосовані інші органы.[ ...]
У ряду риб зяброве дихання на ранніх стадіяхрозвитку не повністю задовольняє потреби організму. Внаслідок цього розвиваються додаткові органи (підкишкова, верхня хвостова та спинна вени), які служать значним доповненням до зябрового дихання. З розвитком і вдосконаленням зябрового дихання ембріональне дихання поступово редукується.
Крім частоти дихання, спостерігаються зміни і в глибині дихання. Риба в деяких випадках (при малому Р02, підвищеній температурі, підвищеному змістіС02 у воді) дуже часто дихає. Самі дихальні рухи невеликі. Таке поверхневе дихання особливо легко спостерігається за підвищеної температури. У деяких випадках риба робить глибоке дихання. Рот і зяброві кришки широко відкриваються і закриваються. При поверхневому диханні дихальний ритм великий, при глибокому - мал.
Спостерігаючи за ритмом дихання риб, М. М. Воскобойніков дійшов висновку, що проходження води в одному напрямку через рот, зяброві пелюстки і зяброві отвори забезпечується роботою зябрових кришок і особливим положенням зябрових пелюсток.
У міру становлення зябрового типу дихання лосось легше використовує кисень, якщо навіть останній знаходиться в малій концентрації (зниження порогової концентрації Ог).
Співвідношення основного дихання та додаткового змінюється у різних риб. Навіть у в'юна кишкове дихання з додаткового перетворилося на майже рівне зябровому дихання. В'юн все ж потребує. кишковому диханні, навіть якщо він знаходиться у добре аерованій воді. Час від часу він піднімається на поверхню та заковтує повітря, а потім знову опускається на дно. Якщо, наприклад, у окуня або коропа при нестачі кисню дихальний ритм частішає, то в'юн в. подібних умовах не частішає ритм дихання, а інтенсивніше використовує кишкове дихання.
Вода прокачується через зяброву порожнину за допомогою руху ротового апарату та зябрових кришок. Тому частоту дихання у риб визначають за кількістю рухів зябрових кришок. На ритм дихання риб в першу чергу впливає вміст у воді кисню, а також концентрація діоксиду вуглецю, температура, pH і т.д. . Наприклад, при 10 °С та нормальному вмісті кисню (4,0-5,0 мг/л) форель здійснює 60-70, короп -30-40 дихальних рухів за хвилину, а при 1,2 мг 02/л частота дихання збільшується у 2-3 рази. Взимку у коропа ритм дихання різко уповільнюється (до 3-4 дихальних рухів за хвилину).
При відкритому роті і закритих зябрових кришках зоду входить у ротову порожнину, проходить між зябровими пелюстками в зяброву порожнину. Це – вдих. Потім рот закривається, а жаберна кришка прочиняється і вода виходить назовні. Це – видих. Розгляд зазначеного процесу в деталях призвело до двох різних уявлень про механізм дихання.
У деяких риб для повітряного дихання пристосовані ковтка і зяброва порожнина.
Зябра є основним органом дихання у більшості риб. Однак можна навести приклади, коли в деяких риб роль зябрового дихання зменшена, а роль інших органів у процесі дихання збільшена. Тому відповісти на питання, чим дихає риба зараз, не завжди можна. Значно розширивши таблицю Бете, ми наводимо співвідношення різних форм дихання у риб у нормальних умовах (табл. 85).
Пригнічуюча дія надлишку С02 на зяброве дихання та стимулювання легеневого дихання у подвійних риб відзначалися неодноразово. Перехід подвійних риб з водного дихання на повітряне супроводжується зниженням артеріального р02 і підвищенням рС02. Особливо слід зазначити, що стимуляція повітряного дихання і пригнічення водного у подвійних риб відбувається під впливом зниження рівня 02 у воді та підвищення рівня С02. Правда, при гіпоксії у подвійних риб ((Чеосегагоскк) посилюється як легеневе, так і зяброве дихання, а при гіперкапнії - тільки легеневе. Цікаво, що при спільній дії гіпоксії та гіперкапнії вентиляція легень зростає, а зябер знижується. в області зябер або у виносних зябрових судинах.
Недорозвиненість або повна відсутність зябрової кришки ускладнює дихання і призводить до захворювання зябер. Косе рило заважає їді. Вигнута спина і мопсовидність голови призводять до значного відставання у рості.
Найбільш поширений тип кишкового дихання той, при якому повітря проганяється через кишечник, і в середній або задній його частині відбувається газообмін (в'юнові, деякі сомові). При іншому типі, наприклад, у Hippostomos і Acarys, повітря після перебування деякого часу в кишечнику не виходить через анус, а видавлюється назад у порожнину рота і потім викидається назовні через зяброві щілини. Цей тип кишкового дихання принципово відрізняється від першого; згодом у деяких риб він розвинувся в легеневе дихання.[ ...]
Більш складне пристосування для повітряного дихання є надджаберний орган. Наджаберний орган є у ОрИ-ocephalus (змієголова), що живе в р. Амур, у Luciocephalus, у Anabas та ін. Цей орган утворений випинання глотки, а не власне зябрової порожнини, як у лабіринтових риб.
Дихальні рухи, дихальний ритм. У риб зяброва кришка періодично відкривається і закривається. Ці ритмічні рухи зябрової кришки давно відомі як дихальні рухи. Однак правильне розуміння процесу дихання було досягнуто порівняно недавно.
Цілком очевидно, що інтенсивність шкірного дихання є вираз пристосованості риби до життя в умовах кисневої недостатності, коли зяброве дихання не в змозі забезпечити організм киснем у потрібній кількості.
Спостерігається загальне правило: з розвитком повітряного дихання відбувається зменшення зябрового (Суворов). Анатомічно це виражається в укороченні зябрових пелюсток (у Polypterus, Ophiocephalus, Arapaima, Electrophorus) або в зникненні цілого ряду пелюсток (у Monopterus, Amphipnous і двоякоїди). У протоптеруса, наприклад, на першій і другій дугах пелюстки майже зовсім відсутні, а у лепідозирена зяброві пелюстки слабо розвинені.
У риб теплих вод є пристрій для повітряного дихання у вигляді лабіринту. Лабіринтовий орган утворений випинанням власне зябрової порожнини і іноді (як у АпаЬаз), забезпечений власною мускулатурою. Внутрішня поверхня «лабіринтової порожнини» має різноманітні викривлення завдяки вигнутим кістковим пластиночкам, покритим слизовою оболонкою. До поверхні «лабіринтової порожнини» підходить багато кровоносних судин та капілярів. Кров до них потрапляє з гілки четвертої зябрової артерії, що приносить. Кров, насичена киснем, йде у спинну аорту. Повітря, яке захоплює риба в рот, потрапляє з порожнини рота в лабіринт і віддає там кисень у кров.
Нещодавно більш ґрунтовні дослідження шкірного дихання на 15 видах риб провела С. В. Стрельцова (1949). Вона визначала як загальне дихання, і спеціально шкірне. Зяброве дихання вимикалося шляхом накладання герметичної гумової маски на зябра. Така методика дозволила їй визначити частку участі шкірного дихання у загальному диханні риби. Виявилося, що ця величина у різних риб дуже різна і пов'язана з способом життя та екологією риб.
Досвідами було показано, що V, VII, IX та X пари головних нервів необхідні для нормального дихання. Гілки від них іннервують верхню щелепу (V пара), зяброву кришку (VII пара) та зябра (IX і X пари).
Пракгатично у всіх круглоротих і риб є «морфофункціональний резерв» підвищення потужності дихання у вигляді деяких «збмгочшх» газообмінних структур. Експериментально встановлено, що в нормальних умовах у риб функціонує не більше 60% зябрових пелюсток. Інші включаються лише в умовах гіпоксії, що настає, або при зростанні потреби в кисні, наприклад при підвищенні швидкості плавання.
У личинковій стадії (пуголовки) земноводні мають велику подібність з рибами: у них зберігається зяброве дихання, є плавці, двокамерне серце і одне коло кровообігу. Для дорослих форм характерні трикамерне серце, два кола кровообігу, дві пари кінцівок. З'являються легкі, але вони розвинені слабо, тому додатковий газообмін відбувається через шкіру (рис. 81). Земноводні мешкають у теплих, вологих місцях, особливо поширені у тропіках, де вони найбільш численні.
Личинок і мальків осетрових риб перевозять у перші два дні після вилуплення з ікри до переходу на зяброве дихання, тому що при зябровому диханні потрібно більше кисню. Насичення води киснем має бути не нижче 30% нормального насичення. При температурі води 14-17 ° С та постійній аерації щільність посадки залежно від маси личинок можна довести до 200 шт. на 1 л води.
У віці 15 діб личинка має збільшені підкишкові «вени, які оплетають кишечник (який уже виконує функцію дихання), і грудний плавець з густо розгалуженими судинами. У віці 57 днів у личинки зовнішні зябра скоротилися і повністю закриті зябрової кришкою. Всі. плавці, крім преанального, добре забезпечені судинами. Ці плавці служать органами дихання (рі£.-67).
У перевірочній ретельно виконаній роботі на тому ж вигляді риб - на струмковій форелі було показано, що вже при pH 5,2 відбувається гіпертрофія слизових клітин зябрового епітелію, а на зябрах накопичується слиз. Надалі зі збільшенням кислотності води до 3,5 відзначено руйнування зябрового епітелію та його відторгнення від опорних клітин. Нагромадження слизу на зябрах у період, коли дихання особливо утруднене, відзначено і в інших видів лососевих риб.
Необхідно збільшити рОг, за якого йде освіта НЬ02. Здебільшого у риб посилюється зяброве дихання та серцевий ритм. При цьому відбувається не тільки підтримка р02 більш високому рівні, а й зниження рСОг. Однак цього організм може досягти тільки в певних межах температури, тому що у водоймищі вода насичена киснем менше при підвищеній температурі, ніж при зниженій. У лабораторних умовах і при перевезеннях живої риби в замкнутих судинах стан риб може бути покращений тим; що з підвищенням температури збільшують ріг у воді штучно, шляхом аерації.
Наджаберний і лабіринтовий органи зустрічаються у змієголов і у тропічних риб (півники, гурамі, макроподи). Вони являють собою мішкоподібні випинання зябрової порожнини (лабіринтовий орган) або глотки (наджаберний орган) і призначені в основному для повітряного дихання.
У європейського горчака судини дихальної мережі досягають більшого розвитку, ніж в інших наших коропових риб. Це результат пристосування організму до життя в зябрової порожнини молюсків на ранніх стадіях розвитку в поганих умовах кисневого режиму. З переходом до життя у воді всі ці пристрої пропадають і залишається тільки розвинене зяброве дихання.
Риби поділяють на хрящових та костистих. Середовищем проживання риб є водоймища, що сформувало особливості їх тіла і створило плавці як органи пересування. Дихання зяброве, а серце двокамерне і одне коло кровообігу.
На думку Р. Ллойда, провідним моментом при цьому є збільшення струму води, що проходить через зябра, і, як наслідок цього, зростання кількості отрути, що досягає поверхні зябрового епітелію з подальшим проникненням в організм. Причому концентрація отрути лежить на поверхні зябрового епітелію визначається як концентрацією отрути у більшості розчину, а й швидкістю дихання. Додамо до цього, що згідно з даними, отриманими М. Шепардом, зі зниженням концентрації кисню у воді вміст гемоглобіну в крові збільшується і, що особливо важливо, збільшується швидкість циркуляції крові через зябра.
Між іншим, тією самою здатністю пояснювали випадки життя груп з зарослим ротом. І тут дослідження показали, що такі кори тягнуть деякий час своє існування, пристосувавши вбирати в себе воду для дихання і разом з нею кілька рачків через зяброві отвори.
Для хордових характерна також наявність нервового пучка у вигляді трубки над хордою та травної трубки під хордою. Далі, для них характерна наявність у зародковому стані або протягом всього життя численних зябрових щілин, що відкриваються назовні з глоткового району травної трубки і є органами дихання. Нарешті, для них характерне розташування серця або судини, що заміщає, на черевній стороні.
Узагальнюючи наявні сьогодні численні експериментальні дані щодо впливу на риб різної екології тривалого чи короткострокового дефіциту кисню, можна зробити низку загальних висновків. Первинна реакція риб на гіпоксію – це посилення дихання за рахунок збільшення його частоти чи глибини. Обсяг зябрової вентиляції при цьому різко зростає. Частота серцевих скорочень падає, ударний об'єм серця збільшується, внаслідок чого об'єм кровотоку залишається незмінним. У процесі розвитку гіпоксії споживання кисню спочатку дещо зростає, потім повертається до норми. При поглибленні гіпоксії ефективність поглинання кисню починає знижуватись, тоді як споживання кисню тканинами підвищується, що створює додаткові труднощі для риб у забезпеченні кисневого запиту в умовах його зниженого вмісту у воді. Напруга кисню в артеріальній та венозній крові, утилізація кисню з води, ефективність його перенесення та ефективність оксигенації крові знижуються.
Запис електрокардіограми проводиться в такий спосіб. Електроди, напаяні на тонкі гнучкі провідники, вводять: один в область серця на вентральній стороні тулуба, а інший - між спинним плавцем і головою на дорсальній стороні. Для запису частоти дихання електроди вводяться в зяброву кришку і рострум. Запис частоти дихання та ритму серцевих скорочень можна вести одночасно через два самостійні жанали електрокардіографа або будь-якого іншого приладу (наприклад, двоканального електроенцефалографа). Риба при цьому може бути як у вільному стані в акваріумі, так і у фіксованому. Запис електрокардіограми можна здійснити лише в умовах повного екранування води акваріума. Екраніровка може бути здійснена двома шляхами: зануренням в.воду пластинки з оцинкованого заліза або припаювання до дна акваріума провідника. Якщо акваріум плексигласовий, його слід встановити на лист заліза.
Порівнюючи ці дані для молоді з даними Купциса для дорослої плітки, легко бачити, що гранична величина у молоді плітки на 49-й день після вилуплення дуже близька до порогової величини для дорослої (відповідно 1 і 0,6-1 мг/л). Отже, після встановлення зябрового дихання здатність використовувати кисень швидко досягає своєї межі.
Істотну роль виведенні надлишку солей грають зябра. Якщо двовалентні іони у значній кількості виводяться через нирки та травний трактто одновалентні (головним чином № і СГ) екскретуються майже виключно через зябра, що виконують у риб подвійну функцію - дихання та виділення. У зябровому епітелії є особливі великі келихоподібні клітини, що містять велику кількість мітохондрій і добре розвинений евдоплазматичний ретикулум. Ці "хлоридні" (або "сольові") клітини розташовані в первинних зябрових пелюстках і на відміну від дихальних клітин пов'язані з судинами венозної системи. Перенесення іонів через зябровий епітелій має характер активного транспорту та йде з витратою енергії. Стимулом екскреторної діяльності хлоридних клітин є підвищення осмолярності крові.
Зважені тверді частинки схильні до утворення нестабільних або стійких суспензій і включають неорганічні, так і органічні компоненти. При підвищенні їх вмісту погіршується пропускання світла, знижується активність фотосинтезу, погіршується зовнішній вигляд води та може порушитися зяброве дихання. У міру того, як тверді частинки осідають на дно, зменшується активність придонної флори та фауни.
В онтогенезі риб спостерігається певна послідовність ролі окремих кисневосприймаючих поверхонь: ікринка севрюги дихає всією поверхнею; у ембріона надходження кисню відбувається переважно через густу мережу капілярів на жовтковому мішку; після виклювання, приблизно на 5-й день, з'являється зяброве дихання, яке потім стає головним.
Вюн піднімається на поверхню води заковтувати повітря при: t = 10 ° 2-3 рази на годину, а при 25-30 ° вже 19 разів. Якщо воду прокип'ятити, тобто зменшити Р02, то в'юн піднімається на поверхню при t = 25-2,7 ° один раз на годину. При t=5° у проточній воді він піднімався на поверхню протягом 8 годин. У цих дослідах досить ясно показано, що кишкове дихання, що є доповненням до зябрового, цілком задовільно справляється зі своєю функцією при малих запитах організму в 02 (при t = 5 °) або при високій концентрації кисню в навколишньому середовищі (проточна вода). Але зябрового дихання недостатньо, якщо обмін в організмі підвищений (t == 25-30 °) або сильно зменшилося Р02 в середовищі (прокип'ячена вода). У цьому випадку додатково включається кишкове дихання, і в'юн отримує необхідну кількість кисню.
У девоні клімат був різко континентальним, посушливим, з різкими коливаннями температури протягом доби та по сезонах, з'явилися великі пустелі та напівпустелі. Спостерігалися й перші заледеніння. У цей період великого розквіту досягли риби, що заселили моря та прісні води. У той час багато наземних водойм висихали в літній період, промерзали зймою, і риб, що їх населяли, могли врятувати два шляхи: заривання в мул або міграція в пошуках води. Першим шляхом пішли двоякодишачі риби, у яких поряд з зябровим розвинулося легеневе дихання (легке розвинулося з плавального міхура). Їхні плавці мали вигляд лопатей, що складаються з окремих кісток з прикріпленими до них м'язами. За допомогою плавців риби могли повзати дном. Крім того, вони також могли мати легеневе дихання. Кістепері риби дали початок першим земноводним – стегоцефалам. На суші в девоні з'являються перші ліси з гігантських папоротей, хвощів та плаунів.
Із загальних клінічних змін у риб відзначаються: пригнічення загального стану, придушення та збочення реакцій на: зовнішні подразнення; потемніння, блідість, гіперемія та крововиливи на шкірних покривах тіла; ерошення луски; порушення почуття рівноваги, орієнтації, координації рухів та узгодженої роботи плавників; кон'юнктивіти, кератити» катаракти, виразки рогівки, витрішкуватість, втрата зору; повна або часткова відмова від прийому корму; припухання черевця (гострі випадки отруєнь); зміна ритму дихання та амплітуди коливання зябрових кришок; періодичні судоми м'язів тулуба, тремор зябрових кришок та грудних плавців. При хронічній інтоксикації розвиваються ознаки наростаючого виснаження. При тяжких процесах розвивається: токсична водянка. При летальному результаті отруєні риби: з води опускаються на дно, вони розвивається кома, дихання стає поверхневим, потім зупиняється - настає смерть.
Менш зрозуміла локалізація периферичних рецепторів, що сприймають зміни у вмісті С02, та шляхи проведення імпульсів з цих рецепторів до дихального центру. Так, наприклад, після перерізання IX і X пар черепних нервів, що іннервують зябра, імпульси зберігалися в ослабленому вигляді. У подвійних риб відмічено пригнічення зябрового дихання зі збільшенням рС02 у воді, яке може бути зняте атропіном. Ефект пригнічення легеневого дихання під впливом надлишку двоокису вуглецю у цих риб не відзначений, що дає підставу припустити наявність у ділянці зябер чутливих до С02 рецепторів.
Клас Земноводні = Амфібії.
Перші наземні хребетні, що ще зберегли зв'язок із водним середовищем. Клас налічує 3900 видів і включає 3 загони: хвостаті (саламандри, тритони), безногі (тропічні черв'яги) та безхвості (жаби, квакші, жаби та ін.).
Вторинно-водні тварини. Оскільки в яйці відсутня амніотична порожнина (разом із круглоротими та рибами амфібії відносяться до анамній), вони розмножуються у воді, де проходять початкові етапи їх розвитку. На різних стадіях життєвого циклу земноводні ведуть наземний або напівводний спосіб життя, поширені практично повсюдно, переважно в районах з високою вологістю на берегах прісних водойм та на сирих ґрунтах. Серед земноводних немає форм, які могли б жити у солоній морській воді. Характерні різні способи пересування: відомі види, що здійснюють досить довгі стрибки, що пересуваються кроком або «повзають», позбавлені кінцівок (черв'яги).
Основні ознаки земноводних.
Амфібії зберегли багато рис своїх суто водних предків, але водночас набули й ряд ознак, властивих справжнім наземним хребетним.
Для хвостатих і безхвостих характерний личинковий розвиток з зябровим диханням у прісній воді (пуголовки жаби) та їх метаморфоз у дорослу особину, що дихає легкими. У безногих після вилуплення личинка набуває вигляду дорослої тварини.
Кровоносна система характеризується двома колами кровообігу. Серце трикамерне. Має один шлуночок і два передсердя.
Відокремлюються шийний та крижовий відділи хребта, що мають по одному хребцю.
Для дорослих земноводних характерні парні кінцівки із шарнірними суглобами. Кінцівки п'ятипалі.
Череп двома потиличними виростками рухомо зчленовується з шийним хребцем.
Тазовий пояс щільно причленовується до поперечних відростків крижового хребця.
Очі мають рухливі повіки і миготливі перетинки для захисту очей від засмічення та висихання. За рахунок опуклої рогівки та сплощеного кришталика покращується акомодація.
Передній мозок збільшується і поділяється на дві півкулі. Середній мозок і мозок розвинені незначно. Від головного мозку відходить 10 пар черепно-мозкових нервів.
Шкіра гола, тобто. позбавлена якихось рогових або кісткових утворень, проникна для води та газів. Тому вона завжди волога - кисень спочатку розчиняється в рідині, що покриває шкіру, після чого дифундує в кров. Те саме відбувається з вуглекислим газом, але у зворотному напрямку.
Нирки, як у риб, первинні = мезонефричні.
Для уловлювання звукових хвиль повітряного середовища з'являється барабанна перетинка, за нею – середнє вухо (барабанна порожнина), в якому розташовується слухова кісточка – стрем'я, що проводить коливання до внутрішнього вуха. Євстахієва труба порожнина середнього вуха повідомляється з ротовою порожниною. З'являються хоани – внутрішні ніздрі, носові ходи стають наскрізними.
Температура тіла непостійна (пойкілотермія) залежить від температури навколишнього середовища і лише трохи перевищує останню.
Ароморфози:
З'явилися легкі та легеневі дихання.
Ускладнилася кровоносна система, розвинувся легеневе коло кровообігу, тобто. у амфібій два кола кровообігу – великий та малий. Серце трикамерне.
Сформувалися парні п'ятипалі кінцівки, що є системою важелів з шарнірними суглобами і призначені для пересування по суші.
Утворився шийний відділ у хребті, що забезпечує рух голови, та крижовий відділ – місце прикріплення тазового пояса.
З'явилися середнє вухо, повіки, хоани.
Диференціювання м'язів.
Прогресивний розвиток нервової системи.
Філогенія.
Земноводні походять від давніх кістеперих риб у девонському періоді палеозойської ери приблизно 350 млн. років тому. Перші земноводні – іхтіостеги – на вигляд нагадували сучасних хвостатих амфібій. У їх будові були риси, властиві рибам, у тому числі рудименти зябрової кришки та органи бічної лінії.
Покров.Двошаровий. Епідерміс багатошаровий, коріум - тонкий, але рясно забезпечений капілярами. У земноводних збереглася здатність виробляти слиз, але з окремими клітинами, як в більшості риб, а оформленими слизовими залозами альвеолярного типу. Крім того, у амфібій часто є зернисті залози з отруйним секретом різного ступеня токсичності. Колір шкіри амфібій залежить від спеціальних клітин – хроматофорів. До них належать меланофори, ліпофори та іридоцити.
Під шкірою у жаб є великі лімфатичні лакуни - резервуари, заповнені тканинною рідиною і дозволяють за несприятливих умов накопичувати запас води.
Скелетпідрозділяється на осьовий та додатковий, як у всіх хребетних. Хребетний стовп більш диференційований на відділи, ніж у риб і складається з чотирьох відділів: шийного, тулубового, крижового та хвостового. Шийний та крижовий відділи мають по одному хребцю. Тулубних хребців у безхвостих зазвичай сім, проте хвостові хребці (приблизно 12) зливаються у єдину кісточку – уростиль. У хвостатих 13 - 62 тулубних та 22 - 36 хвостових хребців; у безногих загальна кількість хребців сягає 200 – 300. Наявність шийного хребця важлива, т.к. на відміну від риб, земноводні не можуть так швидко розгортати своє тіло, а шийний хребець робить голову рухливою, але з малою амплітудою. Амфібії не можуть повертати голову, але можуть нахиляти її.
Хребці у різних амфібій можуть відрізнятися за своїм типом. У безногих і нижчих хвостатих хребці амфіцельні, з хордою, як у риб. У вищих хвостатих хребці опістоцільні, тобто. спереду тіла вигнуті, а ззаду увігнуті. У безхвостих, навпаки, передня поверхня тіл хребців увігнута, а задня вигнута. Такі хребці називаються процільними. Наявність сочленовних поверхонь і сочленовних відростків забезпечує як міцне з'єднання хребців, а й робить осьовий скелет рухливим, що важливо пересування хвостатих амфібій у питній воді без участі кінцівок, з допомогою бічних вигинів тіла. Крім того, можливі вертикальні рухи.
Череп амфібій являє собою видозмінений череп кісткової риби, пристосований до наземного існування. Мозковий череп довічно зберігається переважно хрящовим. Потилична область черепа містить лише дві бічні потиличні кістки, які несуть по сочленовному виростку, за допомогою яких череп кріпиться до хребців. Вісцеральний череп амфібій зазнає найбільших перетворень: виникають вторинні верхні щелепи; утворені міжщелепними (передщелепними) та верхньощелепними кістками. Редукція зябрового дихання призвела до радикальної зміни під'язикової дуги. Під'язична дуга перетворюється на елемент слухового апарату та під'язичну платівку. На відміну від риб, вісцеральний череп амфібій безпосередньо приростає піднебінно-квадратним хрящем на дно мозкового черепа. Такий тип прямої сполуки компонентів черепа без участі елементів під'язичної дуги називається аутостилією. Елементи зябрової кришки у амфібій відсутні.
Додатковий скелет включає кістки поясів і вільних кінцівок. Як і у риб, кістки плечового поясу амфібій знаходяться в товщі м'язів, які пов'язують їх з осьовим скелетом, але пояс не пов'язаний з осьовим скелетом безпосередньо. Пояс забезпечує опору для вільної кінцівки.
Всім наземним тваринам постійно доводиться долати силу важкості, чого не потрібно робити рибам. Вільна кінцівка служить опорою, дозволяє підняти тіло над поверхнею та забезпечує переміщення. Вільні кінцівки складаються з трьох відділів: проксимального (одна кістка), проміжного (дві кістки) та дистального (щодо великої кількості кісток). У представників різних класів наземних хребетних зустрічаються особливості будови тієї чи іншої вільної кінцівки, але вони мають вторинний характер.
У всіх амфібій проксимальний відділ вільної передньої кінцівки представлений плечовою кісткою, проміжний – ліктьовою та променевою кістками у хвостатих та єдиною кісткою передпліччя (вона утворена в результаті зрощення ліктьової та променевої кістки) у безхвостих. Дистальний відділ утворений зап'ястям, п'ястком та фалангами пальців.
Пояс задніх кінцівок зчленовується безпосередньо з осьовим скелетом, з його крижовим відділом. Надійне та жорстке з'єднання тазового пояса з хребетним стовпом забезпечує роботу задніх кінцівок, які більш важливі для переміщення амфібій.
М'язова системавідрізняється від м'язової системи риб. Тулубна мускулатура зберігає метамерну будову лише у безногих. У хвостатих метамерія сегментів порушується, а у безхвостих амфібій починають відокремлюватися ділянки м'язових сегментів, диференціюючись у стрічковоподібні м'язи. Різко збільшується маса мускулатури кінцівок. У риб руху плавців забезпечуються головним чином м'язами, розташованими на тулубі, тоді як п'ятипала кінцівка рухається за рахунок м'язів, розташованої в ній самій. З'являється складна система м'язів – антагоністів – м'язів згиначів та розгиначів. Сегментована мускулатура є лише області хребетного стовпа. Ускладнюється і спеціалізується мускулатура ротової порожнини (жувальна, язика, дна ротової порожнини), яка не тільки бере участь у захопленні та ковтанні їжі, але й забезпечує вентиляцію ротової порожнини та легень.
Порожнина тіла- Загалом. У земноводних у зв'язку із зникненням зябер змінилося відносне положення перикардіальної порожнини. Її виштовхнуло на дно грудної клітки в зону, що прикривається грудиною (або коракоїдом). Над нею в парі цілемічних каналів лежать легені. Порожнини, що вміщають серце та легені. Поділяє плеврокардіальне перетинання. Порожнина, в якій розташовуються легені, повідомляється з головним цілим.
Нервова система.Головний мозок іхтіопсідного типу, тобто. Основним інтегруючим центром є середній мозок, але мозок амфібій має низку прогресивних змін. Мозок амфібій має п'ять відділів і відрізняється від мозку риб, головним чином великим розвитком переднього мозку, повним поділом його півкуль. Крім того, нервова речовина вже вистилає крім дна бічних шлуночків також боки та дах, утворюючи мозковий склепіння – архіпаліум. Розвиток архіпаліуму, що супроводжується посиленням зв'язків з проміжним і особливо середнім мозком, призводить до того, що асоціативна діяльність, що регулює поведінку, здійснюється у земноводних не тільки довгим і середнім мозком, але й півкулями переднього мозку. Подовжені півкулі спереду мають загальну нюхову частку, від якої беруть початок два нюхові нерви. Позаду переднього мозку знаходиться проміжний мозок. На його даху розташовується епіфіз. На нижній стороні мозку є перехрест зорових нервів (хіазму). Від дна проміжного мозку відходить лійка та гіпофіз (нижня мозкова залоза).
Середній мозок представлений у вигляді двох круглих зорових часток. Позаду зорових часток лежить слаборозвинений мозок. Відразу ж за ним розташований довгастий мозок із ромбовидною ямкою (четвертий шлуночок). Довгастий мозок поступово переходить у спинний мозок.
У земноводних від мозку відходить 10 пар головних нервів. Одинадцята пара не розвинена, а дванадцята відходить поза черепа.
Справжніх спинномозкових нервів у жаби 10 пар. Три передні беруть участь в утворенні плечового сплетення, що іннервує передні кінцівки, а чотири задні пари – в утворенні попереково-крижового сплетення, що іннервує задні кінцівки.
Органи чуттязабезпечують орієнтування земноводних у воді та на суші.
Органи бічної лінії є у всіх личинок та у дорослих з водним способом життя. Представлені скупченням чутливих клітин з відповідними нервами, які розкидані по всьому тілу. Чутливі клітини сприймають температурні, болючі, тактильні відчуття, а також зміна вологості та хімічного складу навколишнього середовища.
Органи нюху. У амфібій з кожного боку голови є невелика зовнішня ніздря, яка веде в подовжений мішок, що закінчується внутрішньою ніздрею (хоаною). Хоани відкриваються у передній частині даху ротової порожнини. Спереду від хоан ліворуч і праворуч є по мішку, що відкриваються в носову порожнину. Це т.зв. вомероназальний орган. У ньому багато сенсорних клітин. Його функція полягає у отриманні нюхової інформації про їжу.
Органи зору мають будову, характерну для наземного хребетного. Це виявляється у опуклій формі рогівки, кришталику у вигляді двоопуклої лінзи, у рухливих повіках, що захищають очі від висихання. Але акомодація, як і риб, досягається переміщенням кришталика шляхом скорочення війного м'яза. М'яз розташовується в кільцевому валику, що оточує кришталик, і при її скороченні кришталик жаби дещо висувається вперед.
Орган слуху влаштований за наземним типом. З'являється другий відділ - середнє вухо, в якому міститься слухова кісточка, що вперше з'являється у хребетних - стрем'я. Барабанна порожнина з'єднана з глотковою областю євстахієвої труби.
Поведінка земноводних дуже примітивна, умовні рефлекси виробляються повільно, а швидко згасають. Двигуна спеціалізація рефлексів дуже мала, тому жаба не може сформувати захисний рефлекс відсмикування однієї лапки, і при подразненні однієї кінцівки вона смикає обома лапками.
Травна системапочинається ротовою щілиною, що веде в ротоглоточну порожнину. У ній розташовується мускулиста мова. У неї відкриваються протоки слинних залоз. Мова та слинні залози вперше з'являються у амфібій. Заліза служать тільки для змочування харчової грудки і в хімічній обробці їжі не беруть участь. На міжщелепних, верхньощелепних кістках, сошнику розташовуються прості конічні зуби, які основою прикріплюються до кістки. Травна трубка диференціюється на ротоглоточную порожнину, короткий стравохід, що виконує функцію проведення їжі в шлунок, і шлунок. Пилорическая його частина перетворюється на дванадцятипалу кишку – початок тонкого кишечника. У петлі між шлунком та дванадцятипалою кишкою лежить підшлункова залоза. Тонка кишка плавно переходить у товсту кишку, яка закінчується вираженою прямою кишкою, що відкривається у клоаку.
Травними залозами є печінка з жовчним міхуром та підшлункова залоза. Протоки печінки разом із протокою жовчного міхура відкриваються в дванадцятипалу кишку. Протоки підшлункової залози впадають у протоку жовчного міхура, тобто. самостійного сполучення з кишечником ця залоза немає.
Т.о. система травлення амфібій відрізняється від аналогічної системи риб більшою довжиною травного тракту, кінцевий відділ товстої кишки відкривається в клоаку.
Кровоносна системазамкнута. Два кола кровообігу. Серце трикамерне. Крім того, у серці є венозний синус, що сполучається з правим передсердям, а з правого боку шлуночка відходить артеріальний конус. Від нього відходять три пари судин, гомологічних зябрових артерій риб. Кожна посудина починається самостійним отвором. Усі три судини лівої та правої сторони йдуть спочатку загальним артеріальним стволом, оточеним загальною оболонкою, а потім розгалужуються.
Судини першої пари (вважаючи від голови), гомологічні судин першої пари зябрових артерій риб, називаються сонними артеріями, які несуть кров до голови. По судинах другої пари (гомологічним другий парі зябрових артерій риб) – дугам аорти – кров прямує до задньої частини тіла. Від дуг аорти відходять підключичні артерії, що несуть кров до передніх кінцівок.
По судинах третьої пари, гомологічним четвертій парі зябрових артерій риб - легеневим артеріям - кров прямує в легені. Від кожної легеневої артерії відходить велика шкірна артерія, якою кров прямує у шкіру для окислення.
Венозна кров від переднього кінця тіла збирається по двох парах яремних вен. Остання, зливаючись зі шкірними венами, що вже прийняли підключичні вени, утворює дві передні порожнисті вени. Вони несуть у венозний синус змішану кров, оскільки шкірними венами рухається артеріальна кров.
У личинок земноводних одне коло кровообігу, їхня кровоносна система подібна до кровоносної системи риб.
У амфібій з'являється новий орган кровообігу – червоний кістковий мозок трубчастих кісток. Еритроцити великі, ядерні, лейкоцити неоднакові на вигляд. Є лімфоцити.
Лімфатична система.Крім лімфатичних мішків, розташованих під шкірою, є лімфатичні судини та серця. Одна пара лімфатичних сердець міститься близько третього хребця, інша – поблизу клоачного отвору. Селезінка, що має вигляд невеликого круглого тіла червоного кольору, розташовується на очеревині поблизу початку прямої кишки.
Дихальна система.Важливо відрізняється від дихальної системи риб. У дорослих особин органами дихання є легені та шкіра. Дихальні шляхи через відсутність шийного відділу короткі. Представлені носовою та ротоглотковою порожнинами, а також гортанню. Гортань відкривається безпосередньо у легені двома отворами. Внаслідок редукції ребер легені наповнюються у вигляді заковтування повітря – за принципом нагнітального насоса.
Анатомічно дихальна система амфібій включає ротоглоточную порожнину (верхні повітроносні шляхи) і гортанно - трахеальну порожнину (нижні шляхи), яка безпосередньо переходить в мішковидні легені. Легке у процесі ембріонального розвитку утворюються як сліпий виріст переднього (глоточного) відділу травної трубки, тому й у дорослому стані залишається пов'язаним із глоткою.
Т.о. дихальна система у наземних хребетних анатомічно та функціонально поділяється на два відділи – систему повітроносних шляхів та респіраторний відділ. Повітряні шляхи здійснюють двосторонній транспорт повітря, але не беруть участь у самому газообміні, респіраторний відділ здійснює газообмін між внутрішнім середовищем організму (кров'ю) та атмосферним повітрям. Газообмін відбувається через поверхневу рідину і пасивно йде відповідно до градієнта концентрації.
Система зябрових кришок стає непотрібною, тому зябровий апарат у всіх наземних тварин частково видозмінюється, його скелетні структури частково входять до складу скелета (хрящів) гортані. Вентиляція легень здійснюється з допомогою примусових рухів спеціальної соматичної мускулатури у процесі дихального акта.
Видільна система,як і у риб, представлена первинними або тулубовими нирками. Це компактні тіла червонувато – коричневого кольору, що лежать з обох боків хребта, а не стрічковоподібні, як у риб. Від кожної нирки тягнеться до клоаки тонкий вольфовий канал. У самок жаб він служить тільки сечоводом, а у самців і сечоводом і сім'япроводом. У клоаку вольфові канали відкриваються самостійними отворами. Також окремо відкривається в клоаку та сечовий міхур. Остаточним продуктом азотистого обміну у амфібій є сечовина. У водних личинок амфібій основним продуктом азотистого обміну є аміак, який у вигляді розчину виводиться через зябра та шкіру.
Амфібії - гіперосмотичні тварини по відношенню до прісної води. Внаслідок цього вода постійно надходить до організму через шкіру, яка не має механізмів, що перешкоджають цьому, як у інших наземних хребетних. Морська вода гіперосмотична по відношенню до осмотичного тиску в тканинах земноводних, при поміщенні їх у таке середовище вода через шкіру йтиме з організму. Ось чому земноводні не можуть жити у морській воді та гинуть у ній від зневоднення.
Статева система.У самців органи розмноження представлені парою округлих білуватих насінників, що прилягають до черевної поверхні нирок. Від насінників до бруньок тягнуться тонкі сім'явивідні канальці. Статеві продукти з насінника через ці канальці направляються в тіла нирок, далі в вольфові канали і по них у клоаку. Перед впаданням у клоаку вольфові канали утворюють невелике розширення – насіннєві бульбашки, що служать для тимчасового депонування сперми.
Органи розмноження самок представлені парними яєчниками зернистої будови. Зверху з них розташовуються жирові тіла. Вони накопичуються поживні речовини, які забезпечують формування статевих продуктів під час зимової сплячки. У бічних частинах порожнини тіла розташовані сильно звивисті світлі яйцеводи, або мюллерові канали. Кожен яйцевод у порожнину тіла в області серця відкривається лійкою; нижня маточна частина яйцеводів різко розширена і відкривається у клоаку. Дозрілі яйця через розрив стінок яєчника випадають у порожнину тіла, потім захоплюються воронками яйцеводів і ними переміщаються в клоаку.
Вольфові канали у самок виконують лише функції сечоводів.
У безхвостих земноводних запліднення зовнішнє. Ікринки відразу ж зрошуються насіннєвою рідиною.
Зовнішні статеві ознаки самців:
У самців на внутрішньому пальці передніх кінцівок є статева бородавка, яка досягає особливого розвитку на час розмноження і допомагає самцям утримувати самок при заплідненні ікри.
Самці зазвичай менше самок.
Розвитокземноводних супроводжується метаморфозом. Яйця містять відносно небагато жовтка (мезолецитальні яйця), тому відбувається радіальне дроблення. З яйця виходить личинка – пуголовок, яка за своєю організацією значно ближче до риб, ніж до дорослих амфібій. Він має характерну рибоподібну форму - довгий хвіст, оточений добре розвиненою плавальною перетинкою, з боків голови у нього є дві - три пари зовнішніх перистих зябер, парні кінцівки відсутні; є органи бічної лінії, що функціонує ниркою є пронефрос (припунки). Незабаром зовнішні зябра зникають, а натомість їх розвиваються три пари зябрових щілин з їх зябровими пелюсточками. У цей час схожістю пуголовка з рибою є двокамерне серце, одне коло кровообігу. Потім шляхом випинання з черевної стінки стравоходу розвиваються парні легені. На цій стадії розвитку артеріальна система пуголовка надзвичайно схожа на артеріальну систему кистеперих і подвійних риб, і вся відмінність зводиться до того, що завдяки відсутності четвертої зябра четверта приносить зяброва артерія без перерви переходить в легеневу артерію. Ще пізніше зябра редукуються. Попереду зябрових щілин з кожного боку утворюється складка шкіри, яка поступово розростаючись назад, затягує ці щілини. Пуголовок переходь цілком на легеневе дихання і заковтує ротом повітря. Надалі у пуголовка формується парні кінцівки – спершу передні, потім задні. Однак передні знаходяться довше приховані під шкірою. Хвіст і кишечник починають коротшати, з'являється мезонефрос, личинка поступово переходить від рослинної їжі до тварини і перетворюється на молоду жабу.
У результаті розвитку личинки в неї перебудовуються внутрішні системи: дихальна, кровоносна, видільна, травна. Метаморфоз завершується формуванням мініатюрної копії дорослої особини.
Для амбіста характерна неотенія, тобто. у них розмножуються личинки, яких тривалий час приймали за самостійний вигляд, тому вони мають свою назву – аксолотль. Така личинка більша за дорослу особину. Інший цікавою групою є протеї, які у воді, які протягом життя зберігають зовнішні зябра, тобто. ознаки личинки.
Метаморфоз пуголовка в жабу представляє величезний теоретичний інтерес, т.к. як доводить, що земноводні походять від рибоподібних істот, але дає можливість у подробицях відновити еволюцію окремих систем органів, зокрема систем кровообігу і дихання, під час переходу водних тварин у наземні.
Значенняземноводних полягає в тому, що вони поїдають багатьох шкідливих безхребетних і самі служать їжею іншим організмам у ланцюгах харчування.
У амфібій є два роду дихальних органів (крім шкіри): зябра та легені. Ослаблення зябрового дихання та виступ на сцену легеневого дихання спостерігається вже у Dipnoi; зміни у цьому напрямі спостерігаються у Polypterus, і Lepidosteus. У амфібій зяброве дихання зберігається насамперед у личинок, а потім у тих Уродела, які все життя проводять і у воді (Perennibranchiata в колишніх системах). Зяброві щілини успадковані амфібіями від рибоподібних предків. Зяброві дуги виявлені у стегоцефалу, у личинок і у деяких дорослих (Branchiosauridae). Усі сучасні амфібії у личинковому стані дихають зябрами. Нормально у них закладається 5 вісцеральних мішків та 6-й недорозвинений. Але назовні відкриваються не всі: зябрових щілин утворюється 4 або ще менше. Іноді щілин значно менше, ніж дуг. Наявність щілин та дуг є доказом походження амфібій від риб. Внутрішні зябра, гомологічні зябрам риб, зустрічаються, однак, тільки у личинок Anura у вигляді коротких виростів покривів на дугах, що розділяють зяброві щілини. М'якою зябрової кришечкою (operculum), що розростається з боку гіоїдної дуги, зяброва область прикривається зовні. Зяброві кришечки правої та лівої сторін зливаються між собою з нижньої сторони, залишаючи у деяких Anura парні отвори, а у більшості – одне непарне на лівій стороні тіла.
На ранніх стадіях розвитку у личинок Anura та у всіх інших амфібій є тільки зовнішні зябра, гомологічні, мабуть, зовнішнім зябрам личинок Polypterini та Dipnoi. У Apoda і в Anura зовнішні зябра існують лише в личинковому періоді, на ранніх стадіях розвитку, у Urodela ж, які знову повернулися до водного життя, вони зберігаються все життя. Звідси назва для цих амфібій - постійно зяброві (Perennibranchiata), хоча назва ця, як було сказано, обіймає групи амфібій, що мають різне походження. Зовнішні зябра, ймовірно успадковані амфібіями від кистеперих риб.
Легкі амфібії мають вигляд довгих циліндричних мішків з тонкими стінками (у Urodela) або короткішими (у Anura). У безногих праве легеня розвинене набагато більше лівого. З'явилися легені у предків чотирилапих ще задовго до виходу на сушу. Ми бачимо такі ж легені у подвійних. З'явилися вони, очевидно, як додатковий орган дихання внаслідок недостатнього розвитку зябрового дихання, з одного боку, і можливо несприятливих для дихання умов в водах, що пересихали і псувалися - з іншого. Задня частина зябрової порожнини розвинулася у них додатковий дихальний орган. Спочатку цей орган, що мав вигляд дволопатевого мішка, що відкривався на нижній стороні глотки, був недосконалий: стінки його повинні були бути топкими, хоч і рясно забезпеченими кров'ю, зі слабо розвиненими або майже нерозвиненими перегородками. Як усі зяброві випинання (щілини), він мав гладку вісцеральну мускулатуру і іннервувався блукаючим першим.
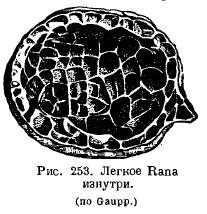
Легкі амфібій мало змінилися у порівнянні з цим: у водних Urodela легені виконують скоріше роль гідростатичного апарату та мають гладку внутрішню поверхню; висота організації їх навіть нижча, ніж у Dipnoi, Нормально ж у амфібій внутрішня поверхня легень є пористою завдяки тому, що в порожнину легені вдається система перекладин (рис. 253). Дуже цікаво, що чим наземнішим є той чи інший вид, тим більш розвиненими є поперечини в легенях: у жаби легше більш комірчасте, ніж жаби. У роду Ascaphus, що живе в гірських струмках, у воді, багатій на кисень, сильно розвинене шкірне дихання, легені ж, навпаки, маленькі і слабо постачаються кров'ю. Ряд амфібій з підряду Salamandroidea (Salamandrina, Plethodon, Spelerpes, Batrachoseps та ін) повністю втратив легені, замість якого глоточное і шкірне дихання сильно розвинулося. .
Легеневі мішки у найпростішому випадку з'єднуються між собою попереду, відкриваючись безпосередньо в глотку поздовжньою щілиною, що підтримується по сторонах хрящовими смужками. Ці хрящові смужки за допомогою м'язів, що прикріплюються до них, можуть розширювати і звужувати гортанну щілину.
Хрящі ці походять з останньої зябрової дуги і в найпростішому вигляді зустрічаються у деяких Urodela. Від цих хрящів можуть відокремлюватися хрящики, які називають персневидними хрящами. Їх можна порівнювати з черпалоподібними хрящами (cartilagines arythenoidea) вищих хребетних. Деякі Urodela, а також Apoda мають досить довге дихальне горло, яке підтримується хрящовими кільцями. У Anura слизова оболонка у гортані утворює голосові зв'язки. Гортань має складну м'язи. На дні або по кутах рота знаходяться резонатори, що надуваються під час квакання.
Механізм дихання наземних амфібій має своїм коренем рефлекси, що спостерігаються у риб та водних амфібій. Найближче до дихання риб дихання личинок Anura, що мають внутрішні зябра, оперкулярну складку та утворену їх зрощенням зяброву порожнину, що відкривається назовні одним отвором. Крім того, у личинок амфібій ротова порожнина рясно забезпечена кров'ю. Забираючи воду в рот і проштовхуючи її шляхом підняття щелеп через ніздрі, личинки збільшують газовий обмін у порожнині рота. Коли ж личинки підростають, вони піднімаються до поверхні, де заковтують повітря подібно до цератоду, і підняттям дна порожнини ротоглотки проштовхують повітря в легені. У водних Urodela спостерігається подібний акт. При опусканні дна ротоглоткової порожнини і при закритих ззаду зябрових отворах вода насмоктується в ротову порожнину через рот або ніздрі або через те й інше разом. Шляхом подальшого підняття дна ротової порожнини при закритих ніздрях вода виштовхується через зяброві щілини. Завдяки цим рухам слизова оболонка рота і глотки вступає в зіткнення з новими масами води, а у зябер виходить рух, що оновлює дихальне середовище.
У наземних амфібій механізм дихання є актом заковтування повітря завдяки опусканню мускулистого дна ротової порожнини і проштовхування їх у легені внаслідок підняття дна. Таким чином, дихання наземних амфібій є актом, що здійснюється на кшталт нагнітального насоса, що переважає у нижчих риб. Безпосередньою основою, де він розвивається, є механізм дихання постоянножаберных амфібій. Цей останній, що спостерігається, наприклад, у Necturus, мав розвинутися у віддалених рибоподібних предків амфібій. З нього вже розвинувся складніший тип дихання наземних - Anura.
У безлегкових саламандр сильно розвинений газообмін внутрішньоротової та глоткової порожнини, що відбувається за допомогою частих, до 120-170 коливань за хвилину діафрагми рота (у жаб їх 30).
Загалом слід сказати, що легеневе дихання в амфібій загалом є допоміжним способом дихання. У цьому також є вказівка і на його філогенетичне походження.
Дихання сучасних амфібій в жодному разі не могло бути джерелом розвитку дихання вищих Tetrapoda (дихання за допомогою підняття ребер, розширення грудної клітки та насмоктування таким чином повітря). Останній тип міг виникнути, у разі намітитися в найдавніших вимерлих амфібій, мали довгі ребра.
Зменшення числа зябер.
Збільшення дихальної поверхні за рахунок утворення зябрових пелюсток.
Освіта зябрових капілярів.
У ланцетника бічні стінки глотки пронизані численними (до 150 пар) косо розташованими зябровими щілинами. До міжзябрових перегородок підходять жаберні артерії, що приносять, а відходять виносять зяброві артерії. Коли вода омиває міжжаберні перегородки, відбувається газообмін між водою, що проходить, і кров'ю, яка тече по тонких судинах перегородок. Зяброві артерії на капіляри не розгалужуються. Крім того, кисень проступає в організм тварини через капіляри шкіри.
У первинноводних хребетних тварин (безщелепні та риби), як і у нижчих хордових, утворюються зяброві щілини, що з'єднують порожнину глотки із зовнішнім середовищем. У круглоротих, з вистилаючої зяброві щілини ентодерми, формуються зяброві мішки (у риб зябра розвиваються з ектодерми). Внутрішня поверхня мішків покрита численними складками – зябровими пелюстками, у стінках яких розгалужується густа мережа капілярів. Мішок внутрішнім вузьким каналом відкривається в глотку (у дорослих мініг - дихальну трубку), а зовнішнім - на бічній поверхні тіла тварини. У міксин від 5 до 16 пар зябрових мішків, у сімейства бделлостомові кожен з них відкривається назовні самостійним отвором, а у сімейства міксинові всі зовнішні зяброві ходи на кожній стороні зливаються в один канал, який відкривається назовні одним отвором, розташованим далеко ззаду. У міног 7 пар зябрових мішків, кожен з яких відкривається назовні самостійним отвором. Дихання здійснюється шляхом ритмічних стисків та розслаблень м'язової стінки зябрової області. У нехарчових мініг вода надходить у дихальну трубку з ротової порожнини, потім омиває пелюстки зябрових мішків, забезпечуючи газообмін, і видаляється через зовнішні зяброві ходи. У круглоротих, що харчуються, вода надходить і виводиться через зовнішні отвори зябрових мішків.
Дихальна система риб має спеціалізовані органи газообміну - ектодермальні зябра, які або розташовані на міжжаберних перегородках, як у хрящових риб, або безпосередньо відходять від зябрових дуг, як у кісткових риб. Обмін газів у зябрах хребетних побудований на кшталт «противоточних систем»: при зустрічному русі кров контактує з багатою киснем водою, що забезпечує її ефективне насичення. Збільшення поверхні поглинання кисню завдяки утворенню зябер супроводжувалося зменшенням числа зябрових щілин у хребетних порівняно з нижчими хордовими. У цільноголових (з хрящових риб) намічається редукція міжжаберних перегородок і утворюється шкіряста зяброва кришка, що прикриває зовні зябра. У кісткових риб у зябрової кришці з'являється кістковий скелет, а міжзяброві перегородки редукуються, що сприяє більш інтенсивному омиванню водою зябрових пелюсток. Поряд з газообміном зябра риб беруть участь у водному та сольовому обміні, у виведенні з організму аміаку та сечовини. Як додаткові органи дихання в окремих груп риб функціонують шкіра, плавальний міхур, надглоткові лабіринти та спеціалізовані ділянки кишкової трубки. У подвійних і багатопероподібних риб з'являються органи повітряного дихання - легені. Легкі виникають як парні вирости черевної частини глотки в області останньої зябрової щілини і коротким каналом пов'язані з стравоходом. Стінки цього виросту тонкі і рясно постачаються кров'ю.
Напрями еволюції легеневого типу дихання
Поява та диференціювання дихальних шляхів.
Диференціювання легені та збільшення дихальної поверхні.
Розвиток допоміжних органів (грудної клітки).
У земноводних у поглинанні кисню та виділенні вуглекислоти беруть участь: у личинок – шкіра, зовнішні та внутрішні зябра, у дорослих – легені, шкіра та слизова оболонка ротоглоткової порожнини. У деяких видів хвостатих земноводних (бузків, протеїв) і у дорослих зберігаються зябра і недорозвиваються або редукуються легені. Співвідношення легеневого та іншого типів газообміну неоднаково: у видів вологих місцепроживання в газообміні домінує шкірне дихання, у мешканців сухих місць більша частина кисню надходить через легені, але шкіра відіграє істотну роль у виділенні вуглекислого газу. Дихальна система дорослих амфібій включає ротоглоточну, гортанно-трахейну порожнини та мішкоподібні легені, стінки яких обплетені густою мережею капілярів. У безхвостих земноводних є загальна гортанно-трахейна камера, у хвостатих вона поділяється на горло і трахею. У гортані з'являються черпалоподібні хрящі, які підтримують її стінку та голосові зв'язки. Легкі хвостатих амфібій являють собою два тонкостінні мішки, що не мають перегородок. У безхвостих усередині легеневих мішків на стінках є перегородки, які збільшують поверхню газообміну (легкі пористі). У земноводних немає ребер, і акт дихання відбувається шляхом нагнітання повітря при вдиху (за рахунок збільшення, а потім зменшення об'єму порожнини ротоглотки) і виштовхування повітря при видиху (за рахунок еластичності стінок легень і черевної мускулатури).
У рептилій відзначається подальше диференціювання дихальних шляхів та значне збільшення функціональної поверхні газообміну у легенях. Повітряні шляхи поділяються на носову порожнину (вона поєднана з ротовою порожниною, але у крокодилів і черепах ці порожнини розділені кістковим небом), горло, трахею і два бронхи. Стінки гортані підтримують парні черпалоподібні та непарні перстоподібні хрящі. У ящірок та змій внутрішні стінки легеневих мішків мають складчасту комірчасту будову. У черепах і крокодилів складна система перегородок вдається у внутрішню порожнину легені настільки глибоко, що легеня набуває губчастої будови. Формується грудна клітина: ребра рухомо з'єднані з хребтом та грудиною, розвиваються міжреберні м'язи. Акт дихання здійснюється завдяки зміні об'єму грудної клітки (реберний тип дихання). У черепах зберігається ротоглотковий тип нагнітання повітря. У водних черепах у воді додатковими органами дихання є багаті капілярами вирости глотки і клоаки (анальні бульбашки). Шкірне дихання у рептилій відсутнє.
У птахів повітроносні шляхи представлені носовою порожниною, гортанню, яка підтримується черпалоподібними та перстнеподібними хрящами, довгою трахеєю та системою бронхів. Легкі невеликі, щільні та мало розтяжні прирощені до ребрів з боків хребетного стовпа. Первинні бронхи утворюються при поділі нижньої частини трахеї і входять у тканину відповідної легені, де розпадаються на 15–20 вторинних бронхів, більшість яких закінчуються сліпо, а частина повідомляється з повітряними мішками. Вторинні бронхи пов'язані між собою дрібнішими парабронхами, від яких відходить безліч тонкостінних пористих бронхіол. Обплетені кровоносними судинами бронхіоли утворюють морфофункціональну структуру легені. З легкими птахів пов'язані повітряні мішки – прозорі еластичні тонкостінні вирости слизової оболонки вторинних бронхів. Об'єм повітряних мішків приблизно в 10 разів перевищує обсяг легень. Вони відіграють дуже важливу роль у здійсненні своєрідного дихального акту птахів: у легені як при вдиху, так і при видиху надходить повітря з високим вмістом кисню – подвійне дихання. Крім інтенсифікації дихання повітряні мішки запобігають перегріву організму при інтенсивному русі. Підвищення внутрішньочеревного тиску при видиху сприяє дефекації. Птахи, що пірнають, збільшуючи тиск у повітряних мішках, можуть зменшувати об'єм і тим самим збільшувати щільність, що полегшує занурення у воду. Шкірне дихання у птахів відсутнє.
У ссавців спостерігається подальше диференціювання дихальних шляхів. Утворюється носова порожнина, носоглотка, вхід у горло прикриває надгортанник (у всіх наземних хребетних крім ссавців гортанна щілина замикається спеціальною мускулатурою), в гортані з'являється щитовидний хрящ, далі йде трахея, яка розгалужується на два бронхи. У легких бронхи багаторазово розгалужуються і закінчуються бронхіолами та альвеолами (число альвеол від 6 до 500 млн.), це значно збільшує дихальну поверхню. Газообмін відбувається в альвеолярних ходах та альвеолах, стінки яких густо обплетені кровоносними судинами. Морфофункціональною одиницею легені ссавців є легеневий ацинус, який утворюється в результаті розгалуження термінальної бронхіоли. Формується грудна клітка, яка відокремлюється діафрагмою від черевної порожнини. Число дихальних рухів від 8 до 200. Дихальні рухи здійснюються двома способами: за рахунок зміни об'єму грудної клітини (реберне дихання) та за рахунок діяльності діафрагмального м'яза (діафрагмальне дихання). У вищих ссавців розвинене шкірне дихання через систему шкірних капілярів, яке грає велику роль газообміні.
Еволюція дихання.
1) Дифузне дихання- Це процес вирівнювання концентрації кисню всередині організму і в навколишньому середовищі. Кисень проникає через оболонку клітини одноклітинних організмів.
2) Шкірне дихання- це обмін газів через шкіру у нижчих черв'яків, у хребетних (риб, амфібій), які мають спеціальні органи дихання.
Зяброве дихання
Перисті зябра(вирости шкіри з обох боків тіла) з'являються у морських кільчастих черв'яків, водних членистоногих, у молюсків у мантійній порожнині.
ЗЯБРИ- органи дихання хребетних тварин, що утворюються як вп'ячування травної трубки.
У ланцетника зяброві щілини пронизують горлянку і відкриваються в околожаберную порожнину з частою зміною води.
У риб зябра з зябрових дуг з зябровими пелюстками, пронизаними капілярами. Вода, що заковтується рибою, потрапляє в ротову порожнину, проходить через зяброві пелюстки назовні, омиває їх і постачає кров киснем.

4) Трахейне та легеневе дихання- ефективніше, оскільки кисень поглинається відразу з повітря, а чи не з води. Характерно для наземних молюсків (мішкоподібні легені), павукоподібних, комах, амфібій, рептилій, птахів, ссавців.
Павукообразнімають легеневі мішки (скорпіони), трахеї (кліщі), а в павуків і те й інше.

НАСЯКІмають трахеї - органи дихання наземних членистоногих - система повітроносних трубочок, що відкриваються дихальними отворами (стигми) на бічних поверхнях грудей та черевця.
АМФІБІЇмають на 2/3 шкірне дихання та на 1/3 легеневе. Вперше з'являються повітроносні шляхи: горло, трахея, зачатки бронхів; легкі – гладкостінні мішки.
РЕПТИЛІЇмають розвинені повітроносні шляхи; легкі комірчасті, шкірного дихання немає.
ПТАХИмають розвинені повітроносні шляхи, легкі губчасті. Частина бронхів розгалужується за межами легень і утворює повітряні мішки.

Повітряні мішки- повітроносні порожнини, з'єднані з дихальною системою, що в 10 разів перевищують обсяг легень, службовці для посилення повітрообміну в польоті, функцію газообміну не виконують. Дихання у спокої здійснюється шляхом зміни об'єму грудної клітки.
Дихання у польоті:
1. При підйомі крил повітря через ніздрі засмоктується в легені та задні повітряні мішки (у легенях I газообмін);
Передні повітряні мішки← легкі - задні повітряні мішки
2. При опусканні крил повітряні мішки стискаються, і повітря із задніх повітряних мішків надходить у легені (у легенях II газообмін).
Передні повітряні мішки - легкі← задні повітряні мішки
Подвійне дихання– це обмін газами у легенях на вдиху та видиху.
ссавці- газообмін майже повністю у легенях (через шкіру та травний канал -2%)

Повітроносні шляхи: носова порожнина → носоглотка → ковтка → гортань → трахея → бронхи (бронхи розгалужуються на бронхіоли, альвеолярні ходи і закінчуються альвеолами - легеневими бульбашками). Легкі губчасті будови складаються з альвеол, обплетених капілярами. Дихальна поверхня збільшена в 50-100 разів у порівнянні з поверхнею тіла. Тип дихання альвеолярний. Діафрагма, що відокремлює грудну порожнину від черевної, а також міжреберні м'язи забезпечують вентиляцію легень. Повний поділ ротової та носової порожнини. Ссавці можуть одночасно дихати і жувати.